6. Biochemistry
Methane oxidizing bacteria oxidize methane using one of two enzyme systems. The particulate methane monooxygenase (pMMO) is a copper-containing enzyme found in nearly all methanotrophs and is a membrane-bound enzyme system (see 6.1). The soluble methane monooxygenase (sMMO) is a different iron-containing enzyme that is found in some but not all methanotrophs. Since the pMMO contains copper, copper acquisition is an important function for these bacteria. MMO activity is usually measured by assaying the production of propylene oxide from propylene using gas chromatography.
|
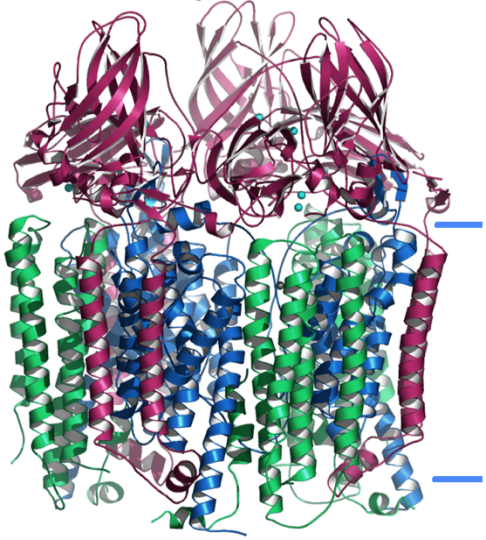
Figure 6.1 Overall structure of the pMMO trimer. The PmoB subunits are shown in magenta, the PmoA subunits are shown in blue, and the PmoC subunits are shown in green. Approximate boundaries of the membrane are shown as blue lines. Copper ions are shown as cyan spheres and zinc ions are shown as gray spheres. |
6.1 pMMO
The membrane-bound or particulate MMO (pMMO) is found in most aerobic methanotrophs, with a few exceptions [Methylocella and Methyloferula strains; (Dedysh et al., 2000; Dedysh et al., 1998; Dunfield et al., 2003)]. The pMMO is a copper-containing enzyme composed of three polypeptides with approximate molecular masses of 45,000 (PmoB ), 26,000 (PmoA) and 23,000 (PmoC) Da (Hakemian and Rosenzweig, 2007; Lieberman and Rosenzweig, 2005a; Myronova et al., 2006). Crystal structures of three pMMOs have been determined (Table 6.1), those from M. capsulatus (Bath) (Lieberman and Rosenzweig, 2005a, b)(Lieberman & Rosenzweig, 2005a, 2005b), M. trichosporium OB3b (Hakemian et al., 2008) (Hakemian et al., 2008), and Methylocystis sp. strain M (Smith et al., 2011)(Smith et al., 2011), with the Methylocystis sp. strain M pMMO structure representing the best quality model (Smith et al., 2011)(Smith et al., 2011). All structures reveal three copies each of the PmoB, PmoA, and PmoC subunits arranged in an a3b3g3 trimer (Fig. 6.1). The PmoB subunit contains both periplasmic and transmembrane domains, while the PmoA and PmoC subunits are composed primarily of transmembrane helices.
Table 1. Structures of pMMO
|
Resolution (Å) |
PDB code |
|
| Methylococcus capsulatus (Bath) pMMO |
2.80 |
1YEW |
| Methylococcus capsulatus (Bath) pMMO |
2.80 |
3RGB* |
| Methylosinus trichosporium OB3b pMMO |
3.90 |
3CHX |
| Methylocystis species strain M pMMO |
2.68 |
3RFR |
*Structure 3RGB is an improved version of structure 1YEW and should be used as the M. capsulatus (Bath) pMMO model; 1YEW is obsolete.
Three metal centers have been modeled in these structures, mononuclear and dinuclear copper sites in the periplasmic domain of PmoB, and a site in the transmembrane region occupied by zinc or copper and coordinated by residues from PmoC. The PmoB monocopper site is only present in the M. capsulatus (Bath) enzyme. The modeling of a dinuclear copper site in PmoB relies on extended X-ray absorption fine structure (EXAFS) data, which indicates the presence of a Cu–Cu interaction at 2.5–2.6 Å (Lieberman et al., 2006; Lieberman et al., 2003)(Lieberman et al., 2003; Lieberman et al., 2006); the crystallographic resolution is not sufficient to distinguish two metal ions, however. Studies employing a soluble fragment of PmoB from M. capsulatus Bath suggest that the active site is located at the crystallographic dicopper site (Balasubramanian et al., 2010; Culpepper et al., 2012).
However, data regarding the number, type, and function of the metal centers associated with the pMMO are not always consistent and some metal-binding ligands are not conserved in all pMMO sequences (Culpepper and Rosenzweig, 2012; Semrau et al., 2010). In addition, isolated pMMO preparations typically have low specific activities, complicating interpretation. Low activities may in part be due to the absence of the physiological reductant, which has yet to be identified. Therefore, uncertainty still exists regarding fundamental aspects of the pMMO structure and mechanism. Additional crystallographic, biochemical, and mechanistic studies are required to resolve these issues (Culpepper and Rosenzweig, 2012; Semrau et al., 2010).
M. capsulatus Bath has been shown to contain an O2-carrier, bacteriohemerythrin, that increases in vitro activity of the pMMO, presumably by providing O2 more efficiently (Chen et al., 2012). A similar gene is found in other methanotroph genome sequences.
6.2 sMMO
|
Figure 6.2 Overall structure of the sMMO dimer. The MmoX subunits are shown in magenta, the MmoY subunits are shown in blue, and the MmoZ subunits are shown in green. The active site iron ions are shown as orange spheres. |
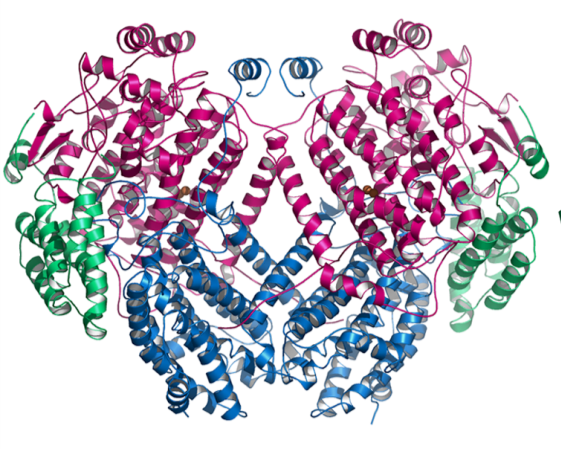
The soluble methane monooxygenase (sMMO) is found in selected methanotrophs (e.g. Methylococcus, Methylosinus, and some genera of Methylocystis, Methylomonas, Methylomicrobium, and Methylovulum). Methylocella silvestris, the most well characterized facultative methanotroph, only contains sMMO (Chen, 2010), as does the obligate methanotroph Methyloferula stellata. Some methanotrophs, such as Methylococcus capsulatus (Bath) and Methylosinus trichosporium OB3b, can express both sMMO and pMMO. In these organisms, sMMO is only expressed under conditions of copper starvation (Murrell et al., 2000; Stanley et al., 1983); the detailed mechanism of this “copper switch” remains unclear. The sMMO system is composed of three components (Merkx et al., 2001). Methane oxidation occurs at a carboxylate-bridged diiron center within the hydroxylase (often referred to as MMOH or H), which has an a2ß2?2 polypeptide arrangement comprising the MmoX, MmoY, and MmoZ subunits (Fig. 6.2) (Dalton, 2005; Rosenzweig et al., 1993). The reductase (MmoC), which contains FAD and a [2Fe-2S] cluster, transfers electrons from NADH to MMOH (Merkx et al., 2001). The third protein, known as the regulatory protein, the effector protein, or protein B (MmoB) is essential for activity and interacts with the hydroxylase, directly affecting access to and from the active site (Lee et al., 2013). All three components have been structurally characterized (Table 6.1), and the diiron active site has been probed by a variety of spectroscopic and computational methods (Merkx et al., 2001; Wallar and Lipscomb, 1996). The mechanism of methane oxidation has been investigated extensively as well. The catalytic cycle begins reaction of the fully reduced diiron(II) center with O2 to form peroxodiiron(III) adducts, which then convert to a diiron(IV) species denoted intermediate Q that is responsible for methane oxidation (Tinberg and Lippard, 2011).
Table 6.1. Structures of sMMO components
| Methylococcus capsulatus Bath MmoXYZ (hydroxylase) | Resolution (Å) | PDB code |
| oxidized 4 °C | 2.20 | 1MMO |
| oxidized | 1.96 | 1FZ1 |
| oxidized | 1.70 | 1MTY |
| reduced in crystal | 2.15 | 1FYZ |
| anaerobically grown reduced | 2.40 | 1FZ5 |
| mixed valence, reduced in crystal | 2.15 | 1FZ2 |
| anaerobically grown mixed valence | 2.07 | 1FZ0 |
| methanol soaked | 2.05 | 1FZ6 |
| ethanol soaked | 1.96 | 1FZ7 |
| Xe pressurized | 3.30 | 1FZI |
| Xe pressurized | 2.60 | 1FZH |
| dibromomethane grown | 2.10 | 1FZ8 |
| iodoethane grown | 2.30 | 1FZ9 |
| pH 8.5 soaked | 2.38 | 1FZ4 |
| pH 6.2 soaked | 2.03 | 1FZ3 |
| Mn(II) soaked | 2.32 | 1XMF |
| apo (metal free) | 2.10 | 1XMG |
| Co(II) reconstituted | 2.32 | 1XMH |
| phenol soaked | 1.96 | 1XU5 |
| 6-bromohexanol soaked | 1.80 | 1XVB |
| 8-bromooctanol soaked | 2.00 | 1XVC |
| 4-fluorophenol soaked | 2.30 | 1XVD |
| 3-bromo-3-butenol soaked | 2.40 | 1XVE |
| chloropropanol soaked | 2.00 | 1XVF |
| bromoethanol soaked | 1.96 | 1XVG |
| bromophenol soaked | 2.30 | 1XU3 |
| Methylosinus trichosporium OB3b MmoXYZ (hydroxylase) | ||
| oxidized | 2.00 | 1MHY |
| oxidized | 2.70 | 1MHZ |
| MmoC, MmoB, and protein-protein complexes | ||
| M. capsulatus Bath MmoB NMR | 2MOB | |
| M. capsulatus Bath MmoC [2Fe-2S] domain NMR | 1JQ4 | |
| M. capsulatus Bath MmoC FAD/NADH binding domain NMR | 1TVC | |
| M. capsulatus Bath MmoXYZ/MmoB complex | 2.90 | 4GAM |
6.3 Copper acquisition
As noted throughout various sections of this website, it is well-known that methanotrophs have a significant response to copper. That is, it has been extensively shown that expression of the two forms of the methane monooxygenase (MMO) is regulated by copper, as is formation of intra-ctyoplasmic membranes and expression of other enzymatic systems including different forms the formaldehyde dehydrogenase.
Given the strong response to copper observed in methanotrophs, it is not surprising that these microbes have multiple systems for the acquisition of copper. Herein we describe the current state-of-knowledge of copper uptake systems in methanotrophs.
6.3.1. MopE or CorA uptake system. At least one methanotroph, Methylococcus capsulatus Bath, secretes a copper regulated, copper-binding protein, MopE, or CorA under copper-limting conditions (Berson and Lidstrom, 1997; Ve et al., 2012). This protein, of 541 amino acids, is found on the cell surface, is also found in a truncated form excreted form, MopE*, that is reported to have a high affinity for copper. As such, it appears that MopE and its modifications may play a role in copper homeostasis in M. capsulatus Bath.
6.3.2. Methanobactin. Most methanotrophs, however, do not have MopE, and instead are found to secrete a small modified polypeptide, or chalkophore for copper acquisition (chalko is Greek for copper, thus chalkophores are analogous to siderophores for iron acquisition). This compound was, first identified in 1993 (Fitch et al., 1993) and then isolated in 1996 (Zahn et al., 1996) and crystallized from Methylosinus. trichosporium OB3b in 2004 (Kim et al., 2004). Subsequent studies have found that this compound, now called methanobactin (mb), has two five-member heterocyclic rings as shown in Fig. 6.3 (Behling et al., 2008). It is these rings, along with associated enethiol groups that are responsible for copper binding, at extremely high affinity of greater than 1021 M-1 (Choi et al., 2006b; El Ghazouani et al., 2011).
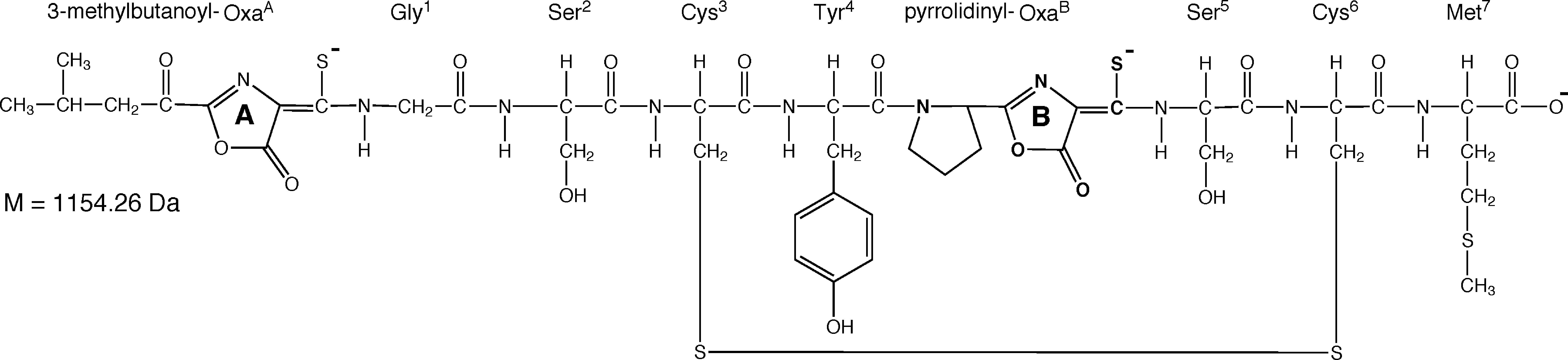
Figure 6.3. Chemical structure of methanobactin from M. trichosporium OB3b
(from Krentz, et al., 2010).
Subsequently, methanobactin has been purified from several other methanotrophs, – Methylocystis strain SB2, Methylocystis hirsuta CSC1, Methylocystis strain M, and Methylocystis rosea (El Ghazouani et al., 2012; Krentz et al., 2010), with some striking variability between the known forms. Specifically, all are small compounds (less than 1500 Da), and all have two heterocyclic rings, one of which is a five-member oxazolone ring, while the other ring is either a five member ring, either oxazolone or imidazolone, or a six-member pyrazinedione ring. Additionally, the rings of all known forms of mb have associated enethiol groups and are separated by two to five amino acids. Finally, all characterized forms of methanobactin bind copper with extremely high affinity, = 1021 M-1 (Bandow et al., 2012; El Ghazouani et al., 2012).
Simple assays have been developed for the screening of microorganisms for the production of methanobactin, based on the Fe-chrome azural S (Fe-CA) assay commonly used for assessment of siderophore production (Schwyn and Neilands, 1987). In this assay, copper is substituted for iron, and multiple references are available for the preparation of this assay (Yoon et al., 2011; Yoon et al., 2010). The assay is simple to perform, either on plates (see Fig. 6.4), or in liquid cultures [please refer to (Yoon et al., 2011) for detailed steps on how to prepare these assays]. Based on these findings, one can then easily purify methanobactin using well-established procedures (Bandow et al., 2011).
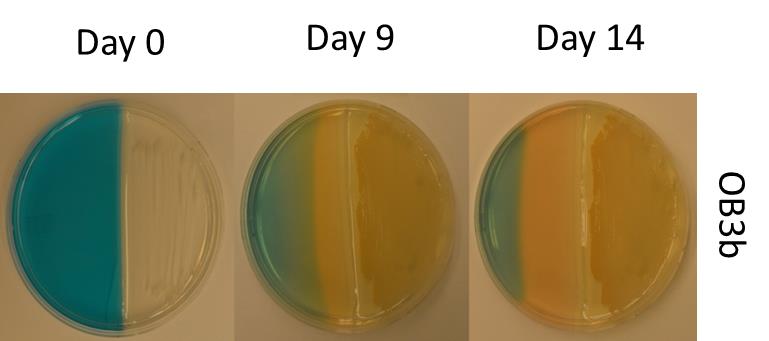
Figure 6.4. Split NMS/50 µM Cu–CAS plates for detection of chalkophore production over time in M. trichosporium OB3b (Semrau et al., 2013). In these assays, methanotrophs are spread on the NMS half as the detergent used to solubilize the Cu-CAS solution inhibits methanotrophic growth. Over time methanobactin production is observed from the decoloration of the Cu-CAS agar (from blue to yellow), indicating that copper has been removed from association with CAS.
Subsequently the molecular biology of methanobactin synthesis has been elucidated, demonstrating that methanobactin from M. trichosporium OB3b is formed from a polypeptide precursor (Semrau et al., 2013), and that this gene is part of a cluster that is replicated in many (but not all) methanotrophs, as well as in some non-methanotrophs (Kenney and Rosenzweig, 2013; Semrau et al., 2013). As such, one can use either chemical screening to determine if a novel microbe expresses mb, or if the genome is available, perform in silico analyses to gauge the ability of novel isolates to produce mb.
6.3.3. Commercial applications of mb
The finding of mb raises some interesting questions as to its role in the canonical copper-switch (Semrau et al., 2013), but it has also been discovered that mb can also bind mercury (Choi et al., 2006a; Vorobev et al., 2013), reducing its toxicity, as a treatment for Wilson disease (Summer et al., 2011; Zischka et al., 2011) as well as also form insoluble gold nanoparticles via the binding and reduction of Au(III) (Choi et al., 2006a). As such, many new areas exist for potential commerical application of methanotrophy.